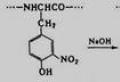
Что такое первичная структура белка кратко. Уровни организации белковых молекул
В полипептидной цепи. Важными особенностями первичной структуры являются консервативные мотивы — сочетания аминокислот, играющих ключевую роль в функциях белка. Консервативные мотивы сохраняются в процессе эволюции видов, по ним часто удаётся предсказать функцию неизвестного белка. Консервати?вные моти?вы — короткие последовательности нуклеотидов в ДНК или аминокислот в белке, которые сохраняются в процессе эволюции, поскольку эти нуклеотиды или аминокислоты незаменимы для выполнения каких-либо процессов в клетке.
Вторичная структура — локальное упорядочивание фрагмента полипептидной цепи, стабилизированное водородными связями. α-спирали — плотные витки вокруг длинной оси молекулы, один виток составляют 3,6 аминокислотных остатка, и шаг спирали составляет 0,54 нм, спираль стабилизирована водородными связями между H и O пептидных групп, отстоящих друг от друга на 4 звена. β-листы (складчатые слои) — несколько зигзагообразных полипептидных цепей, в которых водородные связи образуются между относительно удалёнными друг от друга (0,347 нм на аминокислотный остаток) в первичной структуре аминокислотами или разными цепями белка.
Третичная структура — пространственное строение полипептидной цепи (набор пространственных координат составляющих белок атомов). Структурно состоит из элементов вторичной структуры, стабилизированных различными типами взаимодействий, в которых гидрофобные взаимодействия играют важнейшую роль. В стабилизации третичной структуры принимают участие: ковалентные связи (между двумя остатками цистеина — дисульфидные мостики); ионные связи между противоположно заряженными боковыми группами аминокислотных остатков; водородные связи ; гидрофильно-гидрофобные взаимодействия при взаимодействии с окружающими молекулами воды белковая молекула «стремится» свернуться так, чтобы неполярные боковые группы аминокислот оказались изолированы от водного раствора; на поверхности молекулы оказываются полярные гидрофильные боковые группы.
Четвертичная структура - (субъединичная, доменная) — взаимное расположение нескольких полипептидных цепей в составе единого белкового комплекса. Белковые молекулы, входящие в состав белка с четвертичной структурой, образуются на рибосомах по отдельности и лишь после окончания синтеза образуют общую надмолекулярную структуру.
АК состав белков, пептидная связь и ее физико-хим. хар-ка.
Основными частями и структурными компонентами белковой молекулы являются аминокислоты. В состав пищевых продуктов входит 20 аминокислот, из них 8 не синтезируются в организме человека и являются незаменимыми факторами питания(валин, лейцин, изолейцин, треонин, фенилаланин, триптофан, метионин, лизин). Для детей незаменимой аминокислотой является гистидин. Остальные аминокислоты являются заменимыми, т. е. могут синтезироваться в организме (аланин, аспарагиновая кислота , гликол, глицин , глютаминовая кислота, пролин, серии, тирозин, цистин, цистеин)
Потребность в заменимых аминокислотах удовлетворяется в основном за счет синтеза в организме и частично за счет поступления их в составе пищи. Для изучения аминокислотного состава белков пользуются сочетанием кислотного (НС1), щелочного [Ва(ОН)2] и, реже, ферментативного гидролиза. Гидрофобнае (неполярные, незаряженные): глицин, аланин, валин, лейцин, изолейцин, метионин, пролин, фенилаланин, триптофан. Гидрофильные (полярные, незаряженные): аспарагин, глутамин, серин, треонин, тирозин, цистеин. Гидрофильные (отр. заряд.): аспарагиновая, глутаминовая к-ты. Гидрофильные (пол. заряд.): лизин, аргинин, гистидин.
Пептидная связь — вид амидной связи, возникающей при образовании белков и пептидов в результате взаимодействия α-аминогруппы (—NH 2) одной аминокислоты с α-карбоксильной группой (—СООН) другой аминокислоты. Пептидная связь имеет характеристику частично двойной связи, поэтому она короче, чем остальные связи пептидного остова, и вследствие этого мало подвижна. Электронное строение пептидной связи определяет плоскую жёсткую структуру пептидной группы. Плоскости пептидных групп расположены под углом друг к другу. Пептидные связи очень прочны и самопроизвольно не разрываются при нормальных условиях, существующих в клетках (нейтральная среда, температура тела). В лабораторных условиях гидролиз пептидных связей белков проводят в запаянной ампуле с концентрированной (6 моль/л) соляной кислотой, при температуре более 105 °С, причём полный гидролиз белка до свободных аминокислот проходит примерно за сутки. В живых организмах пептидные связи в белках разрываются с помощью специальных протеолитических ферментов (от англ, protein - белок, lysis - разрушение), называемых также протеазами, или пептидгидролазами.
Цветные реакции на аминокислоты.
Нингидриновая реакция : цветная р-ция на a-аминокислоты, при нагревании (синее окрашивание).
Ксантопротеиновая реакция , жёлтое окрашивание, при кипячении с концентрированной азотной кислотой. После добавления концентрированной щёлочи жёлтое окрашивание переходит в оранжевое. Ароматические АК (фенилаланина, тирозина и триптофана).

Р-ция Адамкевича , триптофан в кислой среде, взаимодействуя с альдегидами кислот, образует продукты конденсации красно-фиолетового цвета.

Р-ция Фоля , Аминокислоты, содержащие сульфгидрильные группы - SH, подвергаются щелочному гидролизу с образованием сульфида натрия Na2S. Последний, взаимодействуя с плюмбитом натрия (образуется в ходе реакции между ацетатом свинца и NaOH), образует осадок сульфида свинца PbS черного или бурого цвета.

Р-ция Миллона , Это реакция на аминокислоту тирозин. Реактив Миллона (раствор HgNO 3 и Hg(NO 2) 2 в разбавленной HNO 3 , содержащей примесь HNO 2) взаимодействует с тирозином с образованием ртутной соли нитропроизводного тирозина, окрашенной в розовато-красный цвет.

Зависимоть конформации белков от их первичной структуры.
В основе каждого белка лежит полипептидная цепь. Она не просто вытянута в пространстве, а организована в трехмерную структуру. Методом рентгеноструктурного анализа Полинг и Кори определили углы пептидных связей, доказав наличие жесткой, планарной(плоской) структуры полипептидной цепи. Несмотря на то что ее конформационная подвижность ограничена, подвижность вокруг одинарных связей при альфа- углеродном атоме возможна. Углы вращения одинарных связей называются торсионными: угол вращения вокруг связи N-C обозначают φ , угол вращения вокруг связи С-С - ψ. Заместители по отношению к пептидной связи могут находится в цис- или транс- положении, причем транс - пептидная связь более стабильна.
Конфигурация - жестко закрепленное ковалентными связями пространственное взаимное расположение частей молекулы (например: принадлежность к L-ряду стереоизомеров или к D-ряду). Для белков также используется понятие Конформация белковой молекулы - определенное, но не застывшее, не неизменное взаимное расположение частей молекулы. Так как конформация белковой молекулы формируется при участии слабых типов связей, то она является подвижной (способной к изменениям), и белок может изменять свою структуру. В зависимости от условий внешней среды молекула может существовать в разных конформационных состояниях, которые легко переходят друг в друга. Энергетически выгодными для реальных условий являются только одно или несколько конформационных состояний, между которыми существует равновесие.
Переходы из одного конформационного состояния в другое обеспечивают функционирование белковой молекулы. Это обратимые конформационные изменения (встречаются в организме, например, при проведении нервного импульса, при переносе кислорода гемоглобином). При изменении конформации часть слабых связей разрушается, и образуются новые связи слабого типа. Фибриллярные белки - это устойчивые, нерастворимые в воде и в разбавленных солевых рас-творах вещества. Располагаясь параллельно друг другу вдоль одной оси, полипептидные цепи образуют длинные волокна (фибриллы) или слои с конформацией b-структуры. Примеры фибриллярных белков: коллаген сухожилий и костной ткани, кератин волос, роговых образований, кожи, ногтей и перьев, эластин упругой соединитель-ной ткани.
Глобу-лярные белки - это соединения, полипептидные цепи которых плотно свернуты в компактные сферические или глобулярные структуры с конформацией a-спирали. Большинство глобу-лярных белков растворимо в водных растворах и легко диффундирует. К ним относятся почти все известные в настоящее время ферменты, а также антитела, некоторые гормоны и многие белки, выполняющие транспортную функцию, например, сывороточный альбумин и гемо-глобин. Некоторые белки принадлежат к промежуточному типу. Подобно фибриллярным белкам, они состоят из длинных, палочковидных структур, и в то же время они, как глобулярные белки, растворимы в водных солевых рас-творах. К таким белкам относятся: миозин - структурный элемент мышц, фибриноген - предшественник фибрина, участвующего в свертывании крови.
Может быть представлено одним из четырех вариантов. Каждый вариант обладает собственными особенностями. Так, существует четвертичная, троичная, вторичная и первичная
Последний в этом списке уровень представляет собой линейную полипептидную цепь из аминокислот. Аминокислоты соединяются друг с другом пептидными связями. Первичная структура белка является простейшим уровнем организации молекулы. Посредством ковалентных пептидных связей между альфа-аминогруппой в одной аминокислоте и альфа-карбоксильной группой в другой обеспечивается высокая стабильность молекулы.
При формировании в клетках пептидных связей активируется сначала карбоксильная группа. После происходит соединение с аминогруппой. Приблизительно так же осуществляется полипептидный лабораторный синтез.
Пептидная связь, представляющая собой повторяющийся фрагмент полипептидной цепи, обладает рядом особенностей. Под воздействием этих особенностей не только формируется первичная структура белка. Они влияют и на высшие организационные уровни полипептидной цепи. Среди основных отличительных черт выделяют копланарность (способность всех атомов, которые входят в пептидную группу, находиться в одной плоскости), трансположение заместителей относительно С-N-связи, свойство существовать в 2-х резонансных формах. К особенностям пептидной связи относят также способность к формированию водородных связей. При этом от каждой пептидной группы может образовываться по две водородные связи с прочими группами (пептидными в том числе). Однако существуют исключения. К ним относят пептидные группы с аминогруппами гидроксипролина или пролина. Они могут формировать только лишь одну Это оказывает воздействие на образование вторичной белковой структуры. Так, на участке, где располагается гидроксипролин или пролин, пептидная цепь легко изгибается, в связи с тем, что нет второй водородной связи, которая удерживала бы ее (как обычно).
Название пептидов формируется из названий аминокислот, входящих в них. Дипептид дают две аминокислоты, трипептид - три, тетрапептид - четыре и так далее. В каждой полипептидной цепи (или пептиде) любой длины присутствует N-концевая аминокислота, в которой содержится свободная аминогруппа, и С-концевая аминокислота, в которой присутствует свободная карбоксильная группа.
Свойства белков.
При изучении этих соединений ученых интересовало несколько вопросов. Исследователи, прежде всего, стремились выяснить размеры, определить форму и массу молекул белков. Следует отметить, что это были достаточно сложные задачи. Трудность состояла в том, что определение по увеличению растворов белков (как это осуществляется у прочих веществ) невозможно, ввиду того, что белковые растворы кипятить нельзя. А определение показателя в соответствии с понижением температуры замерзания результаты дает неточные. Кроме того, белки в чистом виде никогда не встречаются. Однако при помощи разработанных методов было установлено, что колеблется в пределах от 14 до 45 тысяч и больше.
Одной из важных характеристик соединений является фракционное высаливание. Этот процесс представляет собой выделение белков из растворов после прибавления соляных растворов с различными концентрациями.
Еще одним немаловажной характеристикой является денатурация. Этот процесс происходит при осаждении белков тяжелыми металлами. Денатурация представляет собой потерю натуральных свойств. Этот процесс предполагает разные превращения молекулы, кроме разрыва полипептидной цепи. Другими словами, первичная структура белка при денатурации остается неизменной.
П ЕРВИЧНАЯ СТРУКТУРА БЕЛКОВ
Первичная структура белка несет информацию о его пространственной структуре.
1.Аминокислотные остатки в пептидной цепи белков чередуются не случайным образом, а распо-ложены в определенном порядке. Линейная после-довательность аминокислотных остатков в полипеп-тидной цепи называется первичной структурой белка.
2. Первичная структура каждого индивидуально-го белка закодирована в молекуле ДНК (участке, называемом геном) и реализуется в ходе транс-крипции (переписывания информации на мРНК) и трансляции (синтез пептидной цепи).
3. Каждый из 50 000 индивидуальных белков ор-ганизма человека имеет уникальную для данного индивидуального белка первичную структуру. Все молекулы индивидуального белка (например, аль-бумина) имеют одинаковое чередование амино-кислотных остатков, отличающее альбумин от лю-бого другого индивидуального белка.
4. Последовательность аминокислотных остат-ков в пептидной цепи можно рассматривать как
форму запи
си некоторой информации.
Эта информация диктует пространственную ук-ладку длинной линейной пептидной цепи в более компактную трехмерную структуру.
КОНФОРМАЦИЯ БЕЛКОВ
1. Линейные полипептидные цепи индивидуаль-ных белков за счет взаимодействия функциональ-ных групп аминокислот приобретают определен-ную пространственную трехмерную структуру, или конформацию. В глобулярных белках различают
два основных типа конформации
пептидных цепей: вторичную и третичную структуры.
ВТОРИЧНАЯ СТРУКТУРА БЕЛКОВ
2. Вторичная структура белков - это пространст-венная структура, образующаяся в результате взаимодействий между функциональными груп- пами пептидного остова. При этом пептидная цепь может приобретать регулярные структуры двух типов: ос-спирали и р-структуры.
Рис. 1.2. Вторичная структура белка — а-спираль.
В ос-спирали водородные связи образуются между атомом кислорода карбоксильной группы и водородом амидного азота пептидного остова через 4 аминокислоты; боковые цепи аминокислотных остатков располагаются по периферии спирали, не участвуя в образовании водородных связей, фор-мирующих вторичную структуру (рис. 1.2).
Большие объемные остатки или остатки с одина-ковыми отталкивающимися зарядами препятству- ют формированию а-спирали.
Остаток пролина прерывает а-спираль благодаря его кольцевой структуре и невозможности образо-вания водородной связи из-за отсутствия водорода у атома азота в пептидной цепи.
B -Структура формируется между линейными областями одной полипептидной цепи, образуя при этом складки, или между разными полипеп-тидными цепями. Полипептидные цепи или их части могут формировать параллельные (N- и С-концы взаимодействующих пептидных цепей совпадают) или антипараллельные (N- и С-концы взаимодействующих пептидных цепей лежат в противоположных направлениях) р-структуры (рис. 1.3).
В белках также встречаются области с нерегу-лярной вторичной структурой, которые называ-ются беспорядочными клубками, хотя эти структу-ры не так сильно изменяются от одной молекулы белка к другой.
ТРЕТИЧНАЯ СТРУКТУРА БЕЛКОВ
3. Третичная структура белка — это трехмерная пространственная структура, образующаяся за счет взаимодействий между радикалами аминокислот, которые могут располагаться на значительном рас-стоянии друг от друга в пептидной цепи.
Рис. 1.3. Антипараллельная (бета-структура.)
.jpg)
Гидрофобные радикалы аминокислот имеют тенденцию к объединению внутри глобулярной структуры белков с помощью так называемых гид- рофобных взаимодействий и межмолекулярных ван-дер-ваальсовых сил, образуя плотное гидро-фобное ядро. Гидрофильные ионизированные и неионизированные радикалы аминокислот в ос-новном расположены на поверхности белка и оп-ределяют его растворимость в воде.
Гидрофильные аминокислоты, оказавшиеся внут-ри гидрофобного ядра, могут взаимодействовать друг с другом с помощью ионных и водородных свя-зей (рис. 1.4).
.jpg)
Рис. 1.4. Типы связей, возникающие между радикалами аминокислот при формировании третичной структуры белка. 1 — ионная связь; 2 — водородная связь; 3 — гидрофобные взаимодействия; 4 — дисульфидная связь.
|
|
.jpg)
Рис. 1.5. Дисульфидные связи в структуре инсулина человека.
Ионные, водородные и гидрофобные связи отно-сятся к числу слабых: их энергия ненамного пре-вышает энергию теплового движения молекул при комнатной температуре.
Конформация белка поддерживается за счет воз-никновения множества таких слабых связей.
Конформационная лабильность белков — это спо-собность белков к небольшим изменениям кон-формации за счет разрыва одних и образования других слабых связей.
Третичная структура некоторых белков стабили-зирована дисульфидными связями, образующимися за счет взаимодействия SH-групп двух остатков цистеина.
Большинство внутриклеточных белков не имеет ковалентных дисульфидных связей. Их наличие характерно для секретируемых клеткой белков, на-пример дисульфидные связи имеются в молекулах инсулина, иммуноглобулинов.
Инсулин — белковый гормон, синтезирующийся в р-клетках поджелудочной железы. Секретируется клетками в ответ на повышение концентрации глю-козы в крови. В структуре инсулина имеются 2 ди-сульфидные связи, соединяющие 2 полипептидные А- и В-цепи, и 1 дисульфидная связь внутри А-цепи (рис. 1.5).
Особенности вторичной структуры белков ока-зывают влияние на характер межрадикальных вза-имодействий и третичную структуру.
4. Некоторый специфический порядок чередова-ния вторичных структур наблюдается во многих разных по структуре и функциям белках и носит название супервторичной структуры.
Такие упорядоченные структуры часто обозначают как структурные мотивы, которые имеют специфические названия: «а-спираль—поворот—а-спи-раль», «лейциновая застежка-молния», «цинковые пальцы», «структура Р-бочонка» и др.
По наличию а-спиралей и р-структур глобуляр-ные белки могут быть разделены на 4 категории:
1.В первую категорию включены белки, в кото-рых имеются только а-спирали, например миогло-бин и гемоглобин (рис. 1.6).
2. Во вторую категорию включены белки, в кото-рых имеются а-спирали и (3-структуры. При этом а- и (3-структуры часто образуют однотипные со-четания, встречающиеся в разных индивидуаль-ных белках.
Пример. Супервторичная структура типа Р-бочонка.

Фермент триозофосфатизомераза имеет супер-вторичную структуру типа Р-бочонка, где каждая (3-структура расположена внутри р-бочонка и свя-зана с а-спиральным участком полипептидной цепи, находящимся на поверхности молекулы (рис. 1.7, а).
Рис. 1.7. Супервторичная структура типа р-бочонка.
а — триозофосфатизомераза; б — домен пиру ватки назы.
.jpg)
Такая же супервторичная структура обнаружена в одном из доменов молекулы фермента пируваткиназы (рис. 1.7, б). Доменом называют часть молеку-лы, по структуре напоминающую самостоятель-ный глобулярный белок.
Еще один пример формирования супервторич-ной структуры, имеющей Р-структуры и ос-спира-ли. В одном из доменов лактатдегидрогеназы (ЛДГ) и фосфоглицераткиназы в центре располо-жены Р-структуры полипептидной цепи в виде скрученного листа и каждая р-структура связана с а-спиральным участком, расположенным на по-верхности молекулы (рис. 1.8).
Рис. 1.8. Вторичная структура, характерная для многих фер- ментов.
а -домен лактатдегидрогеназы; б— домен фосфоглицераткиназы.
.jpg) 3. В третью категорию включены белки, имею-
щие только вторичную р-структуру. Такие структу-ры обнаружены в иммуноглобулинах, в ферменте супероксиддисмутазе (рис. 1.9).
3. В третью категорию включены белки, имею-
щие только вторичную р-структуру. Такие структу-ры обнаружены в иммуноглобулинах, в ферменте супероксиддисмутазе (рис. 1.9).
Рис. 1.9. Вторичная структура константного домена им-муноглобулина (а)
и фермента супероксиддисмутазы (б).
.jpg)
4. В четвертую категорию включены белки, имеющие в своем составе лишь незначительное ко-личество регулярных вторичных структур. К таким белкам можно отнести небольшие богатые цисти-ном белки или металлопротеины.
В ДНК-связывающих белках имеются общие виды супервторичных структур: «ос-спираль—поворот— ос-спираль», «лейциновая застежка-молния», «цинко- вые пальцы». ДНК-связывающие белки содержат центр связывания, комплементарный участку ДНК с определенной нуклеотидной последовательностью. Эти белки участвуют в регуляции действия генов.
«а-
Спираль—поворот—а-спираль»
Рис. 1.10. Связывание супервторичной
структуры «а-спи-раль—поворот—а-спираль»
в большой бороздке Д
|
|
.jpg)
.jpg) шая бороздка хорошо
при-способлена для связывания белков, имеющих не-большие ос-спиральные участки.
шая бороздка хорошо
при-способлена для связывания белков, имеющих не-большие ос-спиральные участки.
В данный структурный мотив входят 2 ос-спирали: одна более короткая, другая более длинная, соеди-ненные поворотом полипептидной цепи (рис. 1.10).
Более короткая а-спираль располагается попе-рек бороздки ДНК, а более длинная а-спираль на-ходится в большой бороздке, образуя нековалент-ные специфические связи радикалов аминокислот с нуклеотидами ДНК.
Часто белки, имеющие такую структуру, образу-ют димеры, в результате олигомерный белок имеет 2 супервторичные структуры.
Они располагаются на определенном расстоянии друг от друга и выступают над поверхностью белка (рис. 1.11).Две такие структуры могут связываться с ДНК в смежных областях больших бороздок
без значи-тельных изменений в структуре белков.
«Цинковый палец»
«Цинковый палец» — фрагмент белка, содержа-щий около 20 аминокислотных остатков (рис. 1.12).
Атом цинка связан с радикалами 4 аминокислот: 2 остатков цистеина и 2 — гистидина.
В некоторых случаях вместо остатков гистидина находятся остатки цистеина.
Рис. 1.12. Структура участка ДНК-связывающихбелков в форме «цинкового пальца».
.jpg)
Этот участок белка образует а-спираль, которая может специфично связываться с регуляторными участками большой бороздки ДНК.
Специфичность связывания индивидуального регуляторного ДНК-связывающего белка зависит от последовательности аминокислотных остатков, расположенных в области «цинкового пальца».
«Лейциновая застежка-молния»
Взаимодействующие белки имеют а-спиральный участок, содержащий по крайней мере 4 ос-татка лейцина.
Лейциновые остатки расположены через 6 ами-нокислот один от другого.
Так как каждый виток а-спирали содержит 3,6-аминокислотного остатка, радикалы лейцина находятся на поверхности каждого второго витка.
Лейциновые остатки а-спирали одного белка могут взаимодействовать с лейциновыми остатка-ми другого белка (гидрофобные взаимодействия), соединяя их вместе (рис. 1.13).
.jpg) Многие ДНК-связывающие белки взаимодейст-вуют с ДНК в виде олигомерных структур, где субъединицы связываются друг с другом «лейци-новыми застежками». Примером таких белков мо-гут служить гистоны.
Многие ДНК-связывающие белки взаимодейст-вуют с ДНК в виде олигомерных структур, где субъединицы связываются друг с другом «лейци-новыми застежками». Примером таких белков мо-гут служить гистоны.
Гистоны — ядерные белки, в состав которых вхо-дит большое количество положительно заряжен-ных аминокислот — аргинина и лизина (до 80%).
Молекулы гистонов объединяются в олигомер-ные комплексы, содержащие 8 мономеров с по-мощью «лейциновых застежек», несмотря на силь-ный положительный заряд этих молекул.
Резюме. Все молекулы индивидуального белка, имеющие идентичную первичную структуру, при-обретают в растворе одинаковую конформацию.
Таким образом, характер пространственной уклад-ки пептидной цепи определяется аминокислотным составом и чередованием аминокислотных остатков в цепи. Следовательно, конформация — такая же специфическая характеристика индивидуального белка, как и первичная структура.
Выделяют четыре уровня структурной организации белков: первичный, вторичный, третичный и четвертичный. Каждый уровень имеет свои особенности.
Первичной структурой белков называется линейная полипептидная цепь из аминокислот, соединенных между собой пептидными связями. Первичная структура - простейший уровень структурной организации белковой молекулы. Высокую стабильность ей придают ковалентные пептидные связи между α-аминогруппой одной аминокислоты и α-карбоксильной группой другой аминокислоты [показать] .
Если в образовании пептидной связи участвует иминогруппа пролина или гидроксипролина, то она имеет другой вид [показать] .

При образовании пептидных связей в клетках сначала активируется карбоксильная группа одной аминокислоты, а затем она соединяется с аминогруппой другой. Примерно так же проводят лабораторный синтез полипептидов.
Пептидная связь является повторяющимся фрагментом полипептидной цепи. Она имеет ряд особенностей, которые влияют не только на форму первичной структуры, но и на высшие уровни организации полипептидной цепи:


- копланарность - все атомы, входящие в пептидную группу, находятся в одной плоскости;
- способность существовать в двух резонансных формах (кето- или енольной форме);
- транс-положение заместителей по отношению к С-N-связи;
- способность к образованию водородных связей, причем каждая из пептидных групп может образовывать две водородные связи с другими группами, в том числе и пептидными.
Исключение составляют пептидные группы с участием аминогруппы пролина или гидроксипролина. Они способны образовывать только одну водородную связь (см. выше). Это сказывается на формировании вторичной структуры белка. Полипептидная цепь на участке, где находится пролин или гидроксипролин, легко изгибается, так как не удерживается, как обычно, второй водородной связью.
Номенклатура пептидов и полипептидов . Название пептидов складывается из названий входящих в них аминокислот. Две аминокислоты дают дипептид, три - трипептид, четыре - тетрапептид и т. д. Каждый пептид или полипептидная цепь любой длины имеет N-концевую аминокислоту, содержащую свободную аминогруппу, и С-концевую аминокислоту, содержащую свободную карбоксильную группу. Называя полипептиды, перечисляют последовательно все аминокислоты, начиная с N-концевой, заменяя в их названиях, кроме С-концевой, суффикс -ин на -ил (так как аминокислоты в пептидах имеют уже не карбоксильную группу, а карбонильную). Например, название изображенного на рис. 1 трипептида - лейцил фенилаланил треонин .
Особенности первичной структуры белка . В остове полипептидной цепи чередуются жесткие структуры (плоские пептидные группы) с относительно подвижными участками (-СНR), которые способны вращаться вокруг связей. Такие особенности строения полипептидной цепи влияют на укладку ее в пространстве.
Вторичная структура представляет собой способ укладки полипептидной цепи в упорядоченную структуру благодаря образованию водородных связей между пептидными группами одной цепи или смежными полипептидными цепями. По конфигурации вторичные структуры делятся на спиральные (α-спираль) и слоисто-складчатые (β-структура и кросс-β-форма).


α-Спираль . Это разновидность вторичной структуры белка, имеющая вид регулярной спирали, образующейся благодаря межпептидным водородным связям в пределах одной полипептидной цепи. Модель строения α-спирали (рис. 2), учитывающая все свойства пептидной связи, была предложена Полингом и Кори. Основные особенности α-спирали:
- спиральная конфигурация полипептидной цепи, имеющая винтовую симметрию;
- образование водородных связей между пептидными группами каждого первого и четвертого аминокислотных остатков;
- регулярность витков спирали;
- равнозначность всех аминокислотных остатков в α-спирали независимо от строения их боковых радикалов;
- боковые радикалы аминокислот не участвуют в образовании α-спирали.
Внешне α-спираль похожа на слегка растянутую спираль электрической плитки. Регулярность водородных связей между первой и четвертой пептидными группами определяет и регулярность витков полипептидной цепи. Высота одного витка, или шаг α-спирали, равна 0,54 нм; в него входит 3,6 аминокислотных остатка, т. е. каждый аминокислотный остаток перемещается вдоль оси (высота одного аминокислотного остатка) на 0,15 нм (0,54:3,6 = 0,15 нм), что и позволяет говорить о равнозначности всех аминокислотных остатков в α-спирали. Период регулярности α-спирали равен 5 виткам или 18 аминокислотным остаткам; длина одного периода составляет 2,7 нм. Рис. 3. Модель а-спирали Полинга-Кори
β-Структура . Это разновидность вторичной структуры, которая имеет слабо изогнутую конфигурацию полипептидной цепи и формируется с помощью межпептидных водородных связей в пределах отдельных участков одной полипептидной цепи или смежных полипептидных цепей. Ее называют также слоисто-складчатой структурой. Имеются разновидности β-структур. Ограниченные слоистые участки, образуемые одной полипептидной цепью белка, называют кросс-β-формой (короткая β-структура). Водородные связи в кросс-β-форме образуются между пептидными группами петель полипептидной цепи. Другой тип - полная β-структура - характерен для всей полипептидной цепочки, которая имеет вытянутую форму и удерживается межпептидными водородными связями между смежными параллельными полипептидными цепями (рис. 3). Эта структура напоминает меха аккордеона. Причем возможны варианты β-структур: они могут быть образованы параллельными цепями (N-концы полипептидных цепей направлены в одну и ту же сторону) и антипараллельными (N-концы направлены в разные стороны). Боковые радикалы одного слоя помещаются между боковыми радикалами другого слоя.
В белках возможны переходы от α-структур к β-структурам и обратно вследствие перестройки водородных связей. Вместо регулярных межпептидных водородных связей вдоль цепи (благодаря им полипептидная цепь скручивается в спираль) происходит раскручивание спирализованных участков и замыкание водородных связей между вытянутыми фрагментами полипептидных цепей. Такой переход обнаружен в кератине - белке волос. При мытье волос щелочными моющими средствами легко разрушается спиральная структура β-кератина и он переходит в α-кератин (вьющиеся волосы распрямляются).
Разрушение регулярных вторичных структур белков (α-спирали и β-структур) по аналогии с плавлением кристалла называют "плавлением" полипептидов. При этом водородные связи рвутся, и полипептидные цепи принимают форму беспорядочного клубка. Следовательно, стабильность вторичных структур определяется межпептидными водородными связями. Остальные типы связей почти не принимают в этом участия, за исключением дисульфидных связей вдоль полипептидной цепи в местах расположения остатков цистеина. Короткие пептиды благодаря дисульфидным связям замыкаются в циклы. Во многих белках одновременно имеются α-спиральные участки и β-структуры. Природных белков, состоящих на 100% из α-спирали, почти не бывает (исключение составляет парамиозин - мышечный белок, на 96-100% представляющий собой α-спираль), тогда как у синтетических полипептидов 100%-ная спирализация.
Другие белки имеют неодинаковую степень спирализации. Высокая частота α-спиральных структур наблюдается у парамиозина, миоглобина, гемоглобина. Напротив, у трипсина, рибонуклеазы значительная часть полипептидной цепи укладывается в слоистые β-структуры. Белки опорных тканей: кератин (белок волос, шерсти), коллаген (белок сухожилий, кожи), фиброин (белок натурального шелка) имеют β-конфигурацию полипептидных цепей. Разная степень спирализации полипептидных цепей белков говорит о том, что, очевидно, имеются силы, частично нарушающие спирализацию или "ломающие" регулярную укладку полипептидной цепи. Причиной этого является более компактная укладка полипептидной цепи белка в определенном объеме, т. е. в третичную структуру.
Третичная структура белка
Третичной структурой белка называется способ укладки полипептидной цепи в пространстве. По форме третичной структуры белки делятся в основном на глобулярные и фибриллярные. Глобулярные белки чаще всего имеют эллипсовидную форму, а фибриллярные (нитевидные) белки - вытянутую (форма палочки, веретена).
Однако конфигурация третичной структуры белков еще не дает основания думать, что фибриллярные белки имеют только β-структуру, а глобулярные α-спиральные. Есть фибриллярные белки, имеющие спиральную, а не слоисто-складчатую вторичную структуру. Например, α-кератин и парамиозин (белок запирательной мышцы моллюсков), тропомиозины (белки скелетных мышц) относятся к фибриллярным белкам (имеют палочковидную форму), а вторичная структура у них - α-спираль; напротив, в глобулярных белках может быть большое количество β-структур.
Спирализация линейной полипептидной цепи уменьшает ее размеры примерно в 4 раза; а укладка в третичную структуру делает ее в десятки раз более компактной, чем исходная цепь.
Связи, стабилизирующие третичную структуру белка . В стабилизации третичной структуры играют роль связи между боковыми радикалами аминокислот. Эти связи можно разделить на:
- сильные (ковалентные)
[показать]
.
К ковалентным связям относятся дисульфидные связи (-S-S-) между боковыми радикалами цистеинов, находящихся в разных участках полипептидной цепи; изопептидные, или псевдопептидные, - между аминогруппами боковых радикалов лизина, аргинина, а не α-аминогруппами, и СООН-группами боковых радикалов аспарагиновой, глутаминовой и аминолимонной кислот, а не α-карбоксильными группами аминокислот. Отсюда и название этого типа связи - подобная пептидной. Редко встречается эфирная связь, образуемая СООН-группой дикарбоновых аминокислот (аспарагиновой, глутаминовой) и ОН-группой гидроксиаминокислот (серина, треонина).
- слабые (полярные и ван-дер-ваальсовы)
[показать]
.
К полярным связям относятся водородные и ионные. Водородные связи, как обычно, возникают между группой -NН 2 , - ОН или -SН бокового радикала одной аминокислоты и карбоксильной группой другой. Ионные, или электростатические, связи образуются при контакте заряженных групп боковых радикалов -NН + 3 (лизина, аргинина, гистидина) и -СОО - (аспарагиновой и глутаминовой кислот).
Неполярные, или ван-дер-ваальсовы, связи образуются между углеводородными радикалами аминокислот. Гидрофобные радикалы аминокислот аланина, валина, изолейцина, метионина, фенилаланина в водной среде взаимодействуют друг с другом. Слабые ван-дер-ваальсовы связи способствуют формированию гидрофобного ядра из неполярных радикалов внутри белковой глобулы. Чем больше неполярных аминокислот, тем большую роль в укладке полипептидной цепи играют ван-дер-ваальсовы связи.
Многочисленные связи между боковыми радикалами аминокислот определяют пространственную конфигурацию белковой молекулы.
Особенности организации третичной структуры белка . Конформация третичной структуры полипептидной цепи определяется свойствами боковых радикалов входящих в нее аминокислот (которые не оказывают заметного влияния на формирование первичной и вторичной структур) и микроокружением, т. е. средой. При укладке полипептидная цепь белка стремится принять энергетически выгодную форму, характеризующуюся минимумом свободной энергии. Поэтому неполярные R-группы, "избегая" воды, образуют как бы внутреннюю часть третичной структуры белка, где расположена основная часть гидрофобных остатков полипептидной цепи. В центре белковой глобулы почти нет молекул воды. Полярные (гидрофильные) R-группы аминокислоты располагаются снаружи этого гидрофобного ядра и окружены молекулами воды. Полипептидная цепь причудливо изгибается в трехмерном пространстве. При ее изгибах нарушается вторичная спиральная конформация. "Ломается" цепь в слабых точках, где находятся пролин или гидроксипролин, поскольку эти аминокислоты более подвижны в цепи, образуя только одну водородную связь с другими пептидными группами. Другим местом изгиба является глицин, R-группа которого мала (водород). Поэтому R-группы других аминокислот при укладке стремятся занять свободное пространство в месте нахождения глицина. Ряд аминокислот - аланин, лейцин, глутамат, гистидин - способствуют сохранению устойчивых спиральных структур в белке, а такие, как метионин, валин, изолейцин, аспарагиновая кислота, благоприятствуют образованию β-структур. В молекуле белка с третичной конфигурацией встречаются участки в виде α-спиралей (спирализованные), β-структур (слоистые) и беспорядочного клубка. Только правильная пространственная укладка белка делает его активным; нарушение ее приводит к изменению свойств белка и потере биологической активности.
Четвертичная структура белка
Белки, состоящие из одной полипептидной цепи, имеют только третичную структуру. К ним относятся миоглобин - белок мышечной ткани, участвующий в связывании кислорода, ряд ферментов (лизоцим, пепсин, трипсин и т. д.). Однако некоторые белки построены из нескольких полипептидных цепей, каждая из которых имеет третичную структуру. Для таких белков введено понятие четвертичной структуры, которая представляет собой организацию нескольких полипептидных цепей с третичной структурой в единую функциональную молекулу белка. Такой белок с четвертичной структурой называется олигомером, а его полипептидные цепи с третичной структурой - протомерами или субъединицами (рис. 4).

При четвертичном уровне организации белки сохраняют основную конфигурацию третичной структуры (глобулярную или фибриллярную). Например, гемоглобин - белок, имеющий четвертичную структуру, состоит из четырех субъединиц. Каждая из субъединиц - глобулярный белок и в целом гемоглобин тоже имеет глобулярную конфигурацию. Белки волос и шерсти - кератины, относящиеся по третичной структуре к фибриллярным белкам, имеют фибриллярную конформацию и четвертичную структуру.
Стабилизация четвертичной структуры белков . Все белки, у которых обнаружена четвертичная структура, выделены в виде индивидуальных макромолекул, не распадающихся на субъединицы. Контакты между поверхностями субъединиц возможны только за счет полярных групп аминокислотных остатков, поскольку при формировании третичной структуры каждой из полипептидных цепей боковые радикалы неполярных аминокислот (составляющих большую часть всех протеиногенных аминокислот) спрятаны внутри субъединицы. Между их полярными группами образуются многочисленные ионные (солевые), водородные, а в некоторых случаях и дисульфидные связи, которые прочно удерживают субъединицы в виде организованного комплекса. Применение веществ, разрывающих водородные связи, или веществ, восстанавливающих дисульфидные мостики, вызывает дезагрегацию протомеров и разрушение четвертичной структуры белка. В табл. 1 суммированы данные о связях, стабилизирующих разные уровни организации белковой молекулы [показать] .
| Таблица 1. Характеристика связей, участвующих в структурной организации белков | ||
| Уровень организации | Типы связей (по прочности) | Разновидность связи |
| Первичная (линейная полипeптидная цепь) | Ковалентные (сильные) | Пептидная - между α-амино- и α-карбоксильными группами аминокислот |
| Вторичная (α-спираль, β-структуры) | Слабые | Водородные - между пептидными группами (каждой первой и четвертой) одной полипептидной цепи или между пептидными группами смежных полипептидных цепей |
| Ковалентные (сильные) | Дисульфидные - дисульфидные петли в пределах линейного участка полипептидной цепи | |
| Третичная (глобулярная, фибриллярная) | Ковалентные (сильные) | Дисульфидные, изопептидные, сложноэфирные - между боковыми радикалами аминокислот разных участков полипептидной цепи |
| Слабые | Водородные - между боковыми радикалами аминокислот разных участков полипептидной цепи
Ионные (солевые) - между противоположно заряженными группами боковых радикалов аминокислот полипептидной цепи Ван-дер-ваальсовы - между неполярными боковыми радикалами аминокислот полипептидной цепи |
|
| Четвертичная (глобулярная, фибриллярная) | Слабые | Ионные - между противоположно заряженными группами боковых радикалов аминокислот каждой из субъединиц
Водородные - между боковыми радикалами аминокислотных остатков, расположенными на поверхности контактирующих участков субъединиц |
| Ковалентные (сильные) | Дисульфидные - между остатками цистеина каждой из контактирующих поверхностей разных субъединиц | |
Особенности структурной организации некоторых фибриллярных белков
Структурная организация фибриллярных белков имеет ряд особенностей по сравнению с глобулярными белками. Эти особенности можно проследить на примере кератина, фиброина и коллагена. Кератины существуют в α- и β-конформациях. α-Кератины и фиброин имеют слоисто-складчатую вторичную структуру, однако в кератине цепи параллельны, а в фиброине антипараллельны (см. рис. 3); кроме того, в кератине имеются межцепочечные дисульфидные связи, а у фиброина они отсутствуют. Разрыв дисульфидных связей приводит к разъединению полипептидных цепей в кератинах. Напротив, образование максимального числа дисульфидных связей в кератинах путем воздействия окислителей создает прочную пространственную структуру. Вообще у фибриллярных белков в отличие от глобулярных порой трудно строго разграничить разные уровни организации. Если принять (как для глобулярного белка), что третичная структура должна образовываться путем укладки в пространстве одной полипептидной цепи, а четвертичная - нескольких цепей, то в фибриллярных белках уже при формировании вторичной структуры участвует несколько полипептидных цепей. Типичным примером фибриллярного белка является коллаген, который относится к самым распространенным белкам организма человека (около 1/3 от массы всех белков). Он содержится в тканях, обладающих высокой прочностью и малой растяжимостью (кости, сухожилия, кожа, зубы и т. д.). В коллагене треть аминокислотных остатков приходится на глицин, а около четверти или чуть более - на пролин или гидроксипролин.

Изолированная полипептидная цепь коллагена (первичная структура) похожа на ломаную линию. Она содержит около 1000 аминокислот и имеет молекулярную массу порядка 10 5 (рис. 5, а, б). Полипептидная цепь построена из повторяющейся тройки аминокислот (триплет) следующего состава: гли-А-В, где А и В - любые, кроме глицина, аминокислоты (чаше всего пролин и гидроксипролин). Полипептидные цепи коллагена (или α-цепи) при формировании вторичной и третичной структур (рис. 5, в и г) не могут давать типичных α-спиралей, имеющих винтовую симметрию. Этому мешают пролин, гидроксипролин и глицин (антиспиральные аминокислоты). Поэтому три α-цепи образуют как бы скрученные спирали подобно трем нитям, обвивающим цилиндр. Три спиральные α-цепи формируют повторяющуюся структуру коллагена, которая называется тропоколлагеном (рис. 5, г). Тропоколлаген по своей организации является третичной структурой коллагена. Плоские кольца пролина и оксипролина, регулярно чередующиеся вдоль цепи, придают ей жесткость, как и межцепочечные связи между α-цепями тропоколлагена (поэтому коллаген устойчив к растяжению). Тропоколлаген является, по существу, субъединицей фибрилл коллагена. Укладка тропоколлагеновых субъединиц в четвертичную структуру коллагена происходит ступенеобразно (рис. 5, д).
Стабилизация структур коллагена происходит за счет межцепочечных водородных, ионных и ван-дер-ваальсовых связей и небольшого количества ковалентных связей.
α-Цепи коллагена имеют разное химическое строение. Различают α 1 -цепи разных видов (I, II, III, IV) и α 2 -цепи. В зависимости от того, какие α 1 - и α 2 -цепи участвуют в образовании трехцепочечной спирали тропоколлагена, различают четыре типа коллагена:
- первый тип - две α 1 (I) и одна α 2 -цепи;
- второй тип - три α 1 (II)-цепи;
- третий тип - три α 1 (III)-цепи;
- четвертый тип - три α 1 (IV)-цепи.
Наиболее распространен коллаген первого типа: он содержится в костной ткани, коже, сухожилиях; коллаген второго типа содержится в хрящевой ткани и т. д. В одном виде ткани могут быть разные типы коллагена.
Упорядоченная агрегация коллагеновых структур, их жесткость и инертность обеспечивают высокую прочность коллагеновых волокон. Коллагеновые белки содержат также углеводные компоненты, т. е. являются белок-углеводными комплексами.
Коллаген - внеклеточный белок, который образуется клетками соединительной ткани, входящей во все органы. Поэтому с повреждением коллагена (или нарушением его образования) возникают множественные нарушения опорных функций соединительной ткани органов.
| Страница 3 | всего страниц: 7 |
03. Строение белковой молекулы (первичная, вторичная, третичная, четвертичная). Типы связей. Связь структуры с функцией
Каждый белок характеризуется специфической аминокислотной последовательностью и индивидуальной пространственной структурой (конформацией). На долю белков приходится не менее 50% сухой массы органических соединений животной клетки. В организме человека насчитывается до 5 млн. различных видов белков. Белковая молекула может состоять из одной или нескольких цепей, содержащих от пятидесяти до нескольких сотен аминокислотных остатков. Молекулы, содержащие менее пятидесяти остатков, относят к пептидам . В состав многих молекул входят остатки цистеина, дисульфидные связи которых ковалентно связывают участки одной или нескольких цепей. В нативном состоянии белковые макромолекулы обладают специфической конформацией. Характерная для данного белка конформация определяется:
- последовательностью аминокислотных остатков и стабилизируется водородными связями между пептидными и боковыми группами аминокислотных остатков,
- электростатическими и гидрофобными взаимодействиями.
Первичная структура белка. Пептидная связь образуется при реакции аминогруппы одной аминокислоты и карбоксильной группы другой с выделением молекулы воды :
CH3-CH(NH2)-COOH + CH3- CH(NH2)-COOH ^ CH3-CH(NH2)-CO- NH-(CH3) СН-COOH + H2O
Связанные пептидной связью аминокислоты образуют полипептидную цепь . Пептидная связь имеет плоскостную структуру:
- атомы С, О и N находятся в sp -гибридизации;
- у атома N имеется р-орбиталь с неподеленной парой электронов;
- образуется р-п-сопряженная система, приводящая к укорочению связи С-N (0,132 нм) и ограничению вращения (барьер вращения составляет ~63 кДж/моль).
Пептидная связь имеет преимущественно трансконфигурацию относительно плоскости пептидной связи. Подобное строение пептидной связи сказывается на формировании вторичной и третичной структуры белка. Пептидная связь жесткая, ковалентная, генетически детерминированная. В структурных формулах изображается в виде одинарной связи, однако на самом деле эта связь между углеродом и азотом носит характер частично двойной связи. Это вызвано различной электроотрицательностью атомов С, N и O. Вокруг пептидной связи вращение невозможно, все четыре атома лежат в одной плоскости, т.е. компланарны. Вращение же других связей вокруг полипептидного остова достаточно свободно. Последовательность аминокислот для каждого белка уникальна и закреплена генетически.
По числу аминокислотных остатков, входящих в молекулы пептидов, различают дипептиды, трипептиды, тетрапептиды и т.д. Пептиды, содержащие до десяти аминокислотных остатков, называются олигопептидами , содержащие более десяти аминокислотных остатков - полипептидами . Природные полипептиды с молекулярной массой более 6000 называются белками.
Вторичная структура - это пространственное расположение полипептидной цепочки в виде а-спирали или Р-складчатости безотносительно к типам боковых радикалов и их конформации. Л. Полинг и Р. Кори предложили модель вторичной структуры белка в виде а-спирали , в которой водородные связи замыкаются между каждой первой и четвертой аминокислотой, что позволяет сохранять нативную структуру белка, осуществлять простейшие функции, защищать от разрушения. В образовании водородных связей принимают участие все пептидные группы, что обеспечивает максимальную стабильность, снижает гидрофильность и увеличивает гидрофобность белковой молекулы. а-спираль образуется самопроизвольно и является наиболее устойчивой конформацией, отвечающей минимуму свободной энергии. Наиболее распространенным элементом вторичной структуры является правая а-спираль (aR ). Пептидная цепь здесь изгибается винтообразно. Ha каждый виток приходится 3,6 аминокислотного остатка, шаг винта, т.е. минимальное расстояние между двумя эквивалентными точками, составляет 0,54 нм; а-спираль стабилизирована почти линейными водородными связями между NH-группой и СО-группой четвертого по счету аминокислотного остатка. Неполярные или амфифильные а-спирали с 5-6 витками часто обеспечивают заякоривание белков в биологических мембранах (трансмембранные спирали). B складчатых структурах также образуются поперечные межцепочечные водородные связи Если цепи ориентированы в противоположных направлениях, структура называется антипараллельным складчатым листом (ва); если цепи ориентированы в одном направлении, структура называется параллельным складчатым листом (вп). Кроме регулярных в полипептидных цепях есть еще и нерегулярные вторичные структуры, т.е. стандартные структуры, не образующие длинных периодических систем. Это - в-изгибы они называются так потому, что часто стягивают верхушки соседних в-тяжей в антипараллельных в-шпильках). В изгибы обычно входит около половины остатков, не опавших в регулярные структуры белков.
Связи, стабилизирующие третичную структуру :
- электростатические силы притяжения между R-группами, несущими противоположно заряженные ионогенные группы (ионные связи);
- водородные связи между полярными (гидрофильными) R-группами;
- гидрофобные взаимодействия между неполярными (гидрофобными) R-группами;
- дисульфидные связи между радикалами двух молекул цистеина.
Эти связи ковалентные. Они повышают стабильность третичной структуры, но не всегда являются обязательными для правильного скручивания молекулы. В ряде белков они могут вообще отсутствовать.
Третичная структура - уникальное для каждого белка расположение в пространстве полипептидной цепи, зависящее от количества и чередования аминокислот, т.е. предопределенное первичной структурой белка. Конфигурация белковых молекул может быть фибриллярной и глобулярной . Третичная структура многих белков составляется из нескольких компактных глобул, называемых доменами . Между собой домены обычно бывают связаны тонкими перемычками вытянутыми аморфными полипептидными цепями Кроме того, в белках встречаются мотивы укладки полипептидной цепи, похожие на орнаменты на индейских и греческих вазах: мотив меандра, мотив греческого ключа, мотив зигзага-"молнии" При свертывании белковой глобулы значительная часть (не менее половины) гидрофобных радикалов аминокислотных остатков оказывается скрытой от контакта с окружающей белок водой. Происходит образование своеобразных внутримолекулярных «гидрофобных ядер ». В них особенно представлены объемные остатки лейцина, изолейцина, фенилаланина, валина. С появлением третичной структуры у белка появляются новые свойства - биологические . В частности, проявление каталитических свойств связано с наличием у белка третичной структуры. Фибриллярные белки - белки, имеющие вытянутую нитевидную структуру. Большинство фибриллярных белков не растворяется в воде, имеют большую молекулярную массу и высокорегулярную пространственную структуру, которая стабилизируется, главным образом, взаимодействиями (в том числе и ковалентными) между различными полипептидными цепями. Полипептидные цепи многих фибриллярных белков расположены параллельно друг другу вдоль одной оси и образуют длинные волокна (фибриллы) или слои . Глобулярные белки - белки, в молекулах которых полипептидные цепи плотно свёрнуты в компактные шарообразные структуры - глобулы (третичные структуры белка).
Четвертичная структура - это надмолекулярное образование, состоящее из двух и более полипептидных цепей, связанных между собой нековалентно, а водородными связями, электростатическими, дипольдипольные и гидрофобными взаимодействиями между остатками аминокислот, находящихся на поверхности. Каждый из белков-участников третичной структуры при образовании четвертичной структуры называют субъединицей или протомером . Образовавшуюся молекулу называют олигомером, или мультимером . Олигомерные белки чаще построены из четного количества протомеров с одинаковыми или разными молекулярными массами. В образовании четвертичной структуры белка принимают участие те же связи, что и при образовании третичной структуры, за исключением ковалентных. Характерной особенностью белков с четвертичной структурой является их способность к самосборке. Взаимодействие протомеров осуществляется с высокой специфичностью, благодаря образованию десятка слабых связей между контактными поверхностями субъединиц, поэтому ошибки при формировании четвертичной структуры белков исключены.
Практически все белки-ферменты имеют четвертичную структуру и состоят, как правило, из четного числа протомеров (двух, четырех, шести, восьми). Четвертичная структура белка подразумевает такое объединение белков третичной структуры, при котором появляются новые биологические свойства, не характерные для белка в третичной структуре.
Пространственная конфигурация белка т.е. третичная и четвертичная структуры называется конформацией . Если полипептидную цепь взять за концы, растянуть ее и затем отпустить, то она всякий раз будет свертываться в одну и ту же структуру, характерную для этого вида полипептида. В то же время из сказанного, очевидно, следует, что, изменив всего лишь одну аминокислоту в каком-либо полипептиде, мы получим молекулу с совершенно иной структурой, а значит и с иными свойствами.
По химическому составу все белки делят на простые , состоящие только из аминокислотных остатков, и сложные . Сложные могут включать ионы металла (металлопротеины) или пигмент (хромопротеины), образовывать прочные комплексы с липидами (липопротеины), нуклеиновыми кислотами (нуклеопротеины), а также ковалентно связывать остаток фосфорной кислоты (фосфопротеины), углевода (гликопротеины).
Простые белки подразделяют на:
- фибриллярные, растворимые в воде (актин, миозин) и нерастворимые (кератин, эластин, коллаген),
- глобулярные (альбумины, глобулины, протамины, гистоны, проламины).